





















































INTRODUCTION
Marine fish larvae are very vulnerable during the first stages of development and have strict requirements for biotic and abiotic conditions to survive, develop and grow properly (Hamre et al., 2013). The rotifers are now widely used to efficiently farming marine fish larvae and became common in all mass rearing tests. This started from a successful use of rotifers in the mass rearing of red snapper ( Pagrus major ) in Japan (Fujita, 1973, 1979).
The production of sea fish farming is 2,378,328 tons (FAO, 2014), and the number of rotifer required for the production of artificial seeds is up to 7.1E + 11. With the development of the aquaculture industry and the increasing trend of complete culture, the market for live prey is expected to grow even more.
Rotifers seem to have an unbalanced AA profile for marine fish larvae. Rotifers seem deficient in histidine, arginine and lysine for gilthead seabream larvae (Arago et al., 2004c), and unbalanced in histidine, arginine, lysine, threonine and cysteine for both white seabream (Saavedra et al. 2006) and sharpsnout seabream (Saavedra et al. 2007). Histidine is probably the first-limiting AA when rotifers are fed to any of these three Sparid species.
AAs content in live prey (Rotifer,Artemia)
Microalgae (Nannochloropsis gaditana) -enriched rotifer and Artemia had a low relative content of histidine (Fig. 1). That is, histidine is the most limited AA in live preys. Therefore, it is necessary to examine the effect of histidine supplementation on live prey.
.jpg)
Materials and methods (Naz and Turkmen, 2008)
Experimental animals and culture conditions
This study was carried out at the Mediterranean Fisheries Research, Production and Training Institute. Fertilized eggs were collected from broodstock tanks and incubated in conical fiberglass tanks at 16ºC. For the experiment, newly-hatched larvae (approximately 100 larvae/L) were transferred to 300 L fiberglass tanks with black walls. For the first four days after hatching, the temperature was kept at 16ºC0.5ºC, for the following 16 days at 18ºC0.5ºC, and for the final 20 days at 20ºC0.5ºC. Water temperature was controlled by pipe heating systems and automatic transformer equipment was calibrated at 0.5ºC. Salinity was 35-38 g/L throughout the experiment. Oxygen levels were maintained above 6.5 mg/L with liquid oxygen systems. Light (100-150 lux maximum at the water surface) was applied 24 h a day.
Feed regime
All treatment groups were identical for the first 24 days and consisted of Artemia nauplii from day 15 onwards (maximum density 2 prey/mL). On days 25-40, the experimental groups were fed Artemia metanauplii enriched with free histidine (maximum 2 prey/mL) while control larvae were fed unenriched Artemia metanauplii. The investigation was conducted for 16 days to determine the average body weight, protein content, digestive enzymes, and hormones of gilthead seabream (Sparus aurata) larvae fed live prey (Artemia nauplii) enriched with free histidine. The rotifers were reared on bakers yeast and enriched with Selco (INVE Products) prior to transfer to the experimental tanks. Artemia cysts were incubated in continuously aerated and illuminated filtered sea water at 30ºC, collected after 24 h, and washed with tap water. Nauplii were enriched by introducing 600 nauplii/mL into a 15 L round bottom beaker containing 12 L filtered sea water and a 5.3mM FAA solution for 16 h. Gentle aeration ensured homogeneous distribution of the nauplii. Each treatment was conducted in triplicate.
Effects of histidine supplementation on live prey
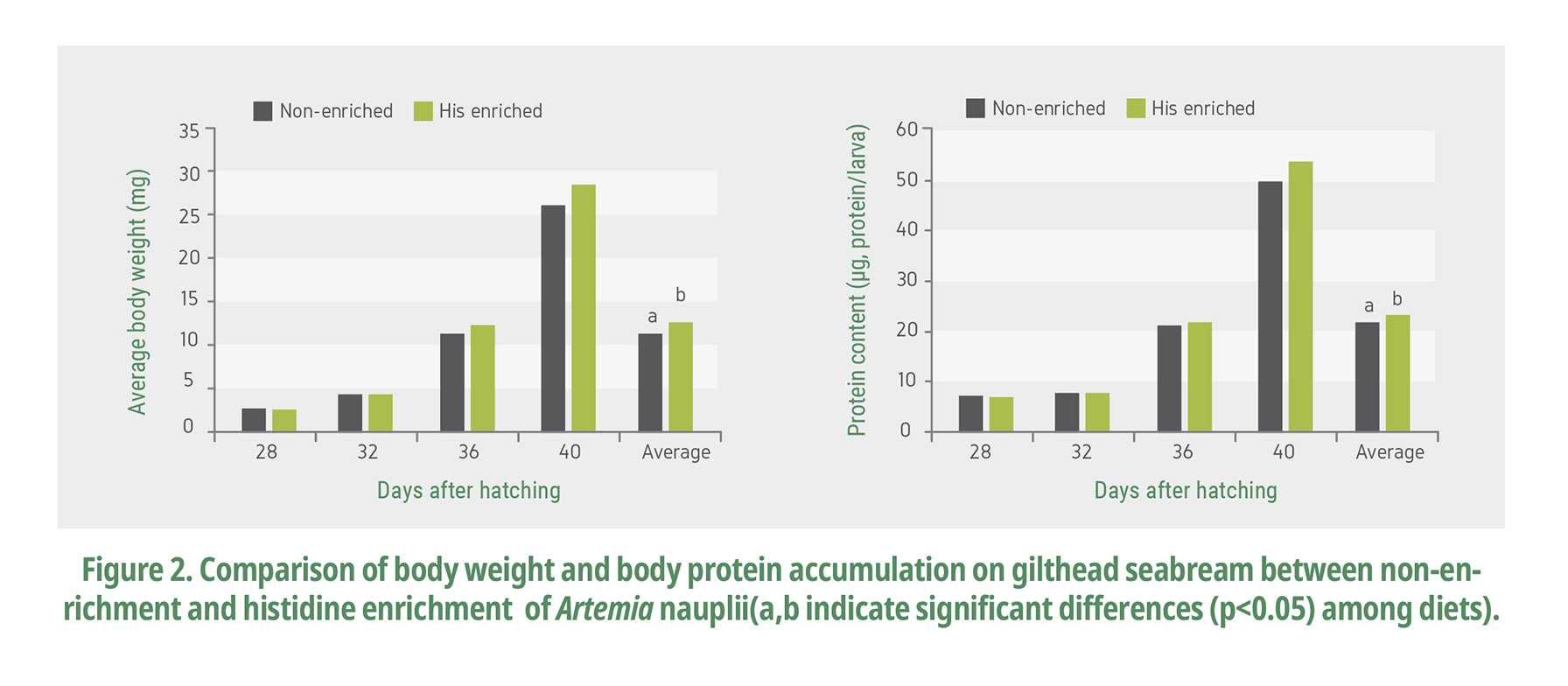
Histidine is the limiting AA for white seabream (Diplodus sargus) larvae (Saavedra et al., 2006). Fig. 2 and 3 show the results of confirming growth performance and digestive related substances level by supplying free histidine-rich Artemia to gilthead seabream larvae. Larvae fed with the enriched Artemia had significantly higher (p<0.05) body weight, protein contents than the control (Fig 2). On day 40, after 16 days of consuming histidine-enriched Artemia, the average weight of the enriched groups was higher than that of the control, indicating that enriching Artemia nauplii with free histidine has a positive effect on seabream growth (Naz and Turkmen, 2008).

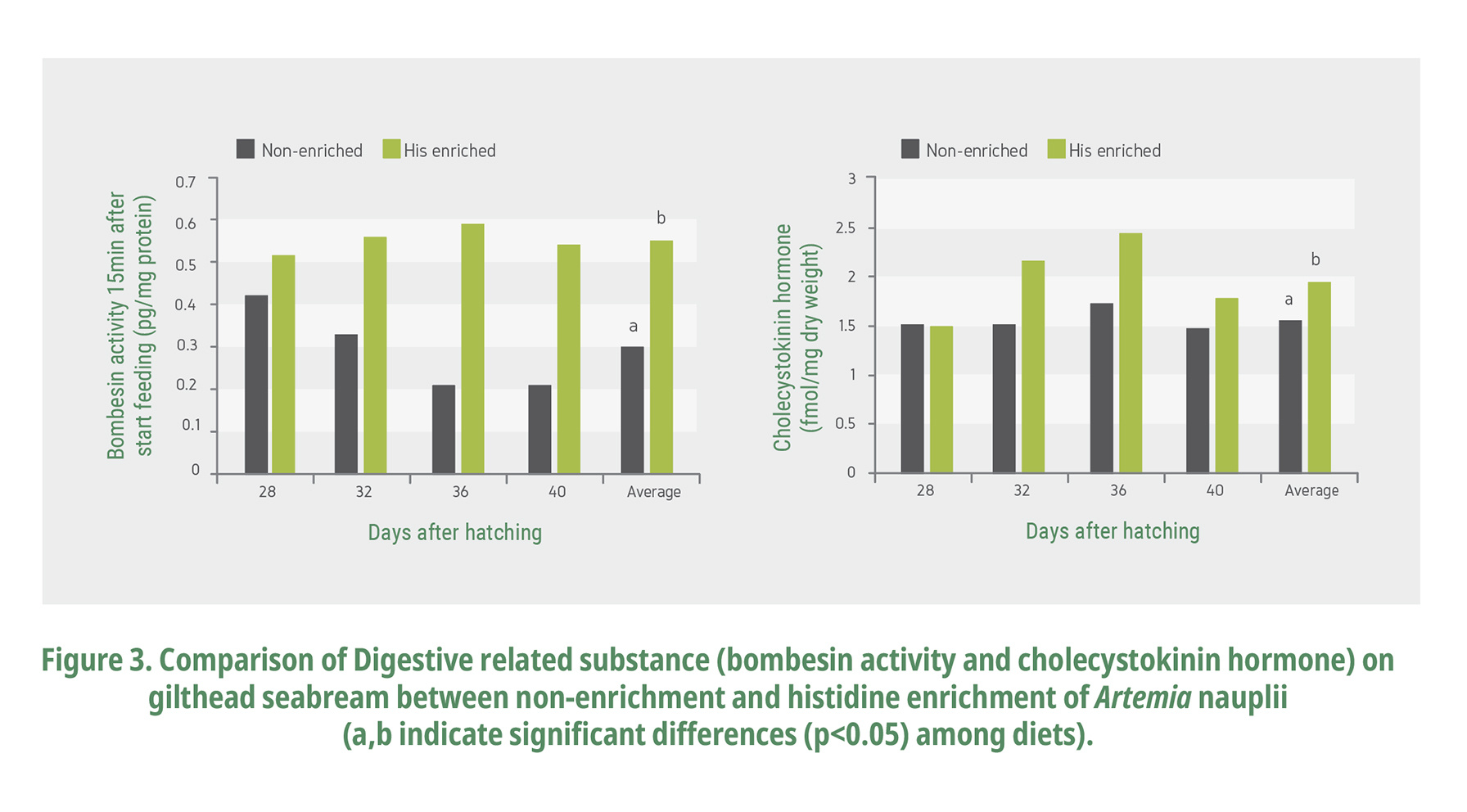
The pituitary neuropeptides, bombesin (gastrin-releasing peptide) and cholecystokinin (CCK), are integral parts of this gastro-entero-pancreatic endocrine system (Moons et al., 1992; Himick and Peter, 1994ab). Bombesin is one of neurotic peptide that stimulates gastric acid secretion, gallbladder contraction, and cholestasis. It influences digestion by activating the peristaltic movement of the gut and the release of HCl as well as increasing blood circulation to the gut wall (McDonald et al., 1979). Cholecystokinin is a kind of polypeptide, promoting pancreas enzyme secretion. Cholecystokinin stimulates gallbladder contraction and secretion of pancreatic enzymes including trypsinogen, the precursor of the key proteolytic enzyme, trypsin (Vander et al., 1998).
Bombesin activity and CCK levels tended to be higher in the larvae fed with the enriched Artemia. There were significant differences in the bombesin level between the enriched group and the control (Fig. 3). Digestive end-products such as L-lysine can act directly on pancreatic acinar cells to stimulate enzyme secretion (Grendell and Rothman, 1981) and FAA can stimulate CCK secretion (Liddle, 2000). Atlantic herring larvae, tube-fed a physiological saline, did not display a CCK response despite the presence of peristaltic movements and apparent fullness of the gut (Koven et al., 2002), indicating that distension of the gut wall is not a trigger for CCK synthesis. Likewise, these results on day 40 reveal that CCK was triggered by the histidine enrichment of the Artemia. The mechanism of histidine-induced digestive enzyme secretion (stimulation) reaction is unclear, but it is thought that digestive enzymes with increased secretion will give positive stimulation to the growth of fish larvae with less digestive absorption capacity. And this evidence reflects the growth of the fish larvae in this experiment (protein accumulation).
CONCLUSION
Artemia nauplii can successfully be enriched with free histidine. Also, bombesin and CCK activity can be stimulated by free histidine (Naz and Turkmen, 2008). As such, histidine is important for enhancing the nutrition of live prey for marine fish larvae, and more research seems to be needed.
REFERENCES
MAIN REFERENCE :
Naz and Turkmen, 2008. Digestive enzymes and hormones in gilthead seabream larvae (Sparus aurata) fed Artemia nauplii enriched with free histidine. The Israeli Journal of Aquaculture Bamidgeh 60(4):230-236.
1. Arago et al., 2004a. A balanced dietary amino acid profile improves amino acid retention in post-larval Senegalese sole (Solea senegalensis). Aquaculture 233:293304
2. Arago et al., 2004b. Amino acid pools of rotifers and Artemia under different conditions: Nutritional implications for fish larvae. Aquaculture 234:429445
3. Arago et al., 2004c. Estimated amino acid requirements during early ontogeny in fish with different life styles: gilthead seabream (Sparus aurata) and Senegalese sole (Solea senegalensis). Aquaculture 242:589605
4. Fujita, 1973. Importance of zooplankton mass culture in producing marine fish seed for fish farming. Bull. Plankton Soc. Jpn. 20:49-53.
5. Fujita, 1979. Culture of red sea bream, Pagrus major, and its food. Cultivation of fish fry and its live food. Spec. Publ. Europ. Maricult. Soc. 4:183-197
6. Grendell and Rothman, 1981. Digestive end products mobilize secretory proteins from subcellular stores in the pancreas. Am. J. Physiol. 241:G67-G73
7. Hamre et al., 2013. Fish larval nutrition and feed formulation: knowledge gaps and bottlenecks for advances in larval rearing. Reviews in Aquaculture 5(1):S26S58
8. Himick and Peter, 1994a. Bombesin acts to suppress feeding behavior and alter serum growth hormone in gold fish. Physiol. Behav. 55:65-72
9. Himick and Peter, 1994b. CCK/gastrin-like immunoreactivity in brain and gut, and CCK suppression of feeding in gold fish. Am. J. Physiol. 267:841-851
10. Koven et al., 2002. The stimulatory effect of ingested protein and/or free amino acids on the secretion of the gastro-endocrine hormone, cholecystokinin (CCK) and the protease, trypsin, in first feeding herring larvae, Clupea harengus. Mar. Biol. 140:1241-1247
11. Liddle. 1995. Regulation of cholecystokinin secretion by intraluminal-releasing factors. Am. J. Physiol., 269:319-327
12. Lubzens et al., 1989. Rotifers as food in aquaculture. Hydrobiologia 186/187:387-400
13. McDonald et al., 1979. Characterization of gastrin releasing peptide from porcine non-antral gastric tissue. Biochem. Biophys. Res. Commun. 90:227
14. Moons et al., 1992. Comparative distribution of substance P (SP) and cholecystokinin (CCK) binding sites and immunorectivity in the brain of the sea bass (Dicentrarchus labrax). Gen. Comp. Endocrinol. 73:270-283
15. Saavedra et al., 2006. Amino acid profiles of Diplodus sargus (L., 1758) larvae: Implications for feed formulation. Aquaculture 261:587593
16. Saavedra et al., 2007. Evaluation of bioavailability of individual amino acids in Diplodus puntazzo larvae: Towards the ideal dietary amino acid profile. Aquaculture 263:192198
17. Vander et al., 1998. The digestion and absorption of food. In: A. Vander, J. Sherman, D. Luciano (eds.). Human Physiology, the Mechanism of Body Function. McGraw Hill, Boston.